本文是Brent Mishler 和 Bruce Baldwin2021年发表在美国加利福尼亚州本地植物协会季刊《ARTEMISIA》上的研究文章。

加利福尼亚州是美国植物种类最为丰富的州之一,拥有大约 6000 种本地植物,其中有超过 40% 的物种是特有的,即只分布于该州。加利福尼亚州地理位置靠近太平洋,气候温暖湿润,同时又有高山和沙漠等不同的地形地貌,这些因素使得该州的植物种类和分布非常丰富,植物生态多样性非常高,包括沙漠、草原、森林、湿地、海岸等多种生态系统。加利福尼亚州拥有许多特有物种,例如红杉、加州野玫瑰、加州橡树、加州石松等,是世界重要的生物多样性重点地区之一。
加利福尼亚州本地植物协会成立于1965年,一直是植物科学和本土植物研究领域的领军人物,是世界上最重要的本土植物组织之一。其长期出色的工作形成了大量植物研究数据,其建立的Calscape数据库涵盖了7900多种加利福尼亚原生植物,除了植物的详细信息外,还利用谷歌地图建立了植物地理空间分布数据,开拓了从植物空间分布上开展研究的领域。
本文提出了在植物“空间系统发育”的角度去思考生物多样性保护的决策。要充分认识到植物分类的维度不仅仅是传统的形态分类,通过基因组数据和空间进行植物分类研究,是发现生命之树进化规律和物种分类新维度的一个重要手段,一个物种可能会存在不同的分类维度。在通过植物移植等手段来开展生物多样性保护时,要考虑原生基因的影响和变异。

正文:
早在达尔文革命之前,物种就被普遍理解为是构成生物多样性总和的离散分布。尽管现在已收集了大量的物种遗传和基因组数据,并伴随了进化理论和研究方法的进步,但生物多样性仍然几乎是通过亚里士多德的视角来看待的,简单地说就是对存在的物种数量的计数。但我们现在知道,生物多样性远不止是一个物种数量的问题——在传统命名的物种之间存在着显著的差异,体现出它们之间存在着重要的、不同的系统发育关系。
(亚里士多德在他的著作《动物学》中提出了一种系统的生物分类方法。这一方法被认为是古希腊分类学的高峰,也成为了现代生物学分类学的基础。亚里士多德的生物分类学方法基于动物和植物的相似性和区别,将它们按照一定的分类方式进行分组。他首先将生物按照其不同的生活方式和行为方式进行分类,然后将它们按照其共性和差异性进行分类,最终建立了一种系统的分类方法。亚里士多德的生物分类学方法强调生物之间的联系和相互关系,这一方法反映了他对于生物多样性的深刻理解和认识。现代生物学家通过对生物的形态、结构、遗传等方面的研究,建立了一种基于亚里士多德分类学方法的生物分类系统。这一分类系统将生物按照其形态、结构、遗传等方面的相似性和差异性进行分组,从而更好地了解生物之间的相互关系和联系。)

物种的名称并不代表物种之间的差异或相似,它只代表了以人类角度去识别的生物(即生命之树)系统的一个“断面”,但它们之间的寿命、形态和遗传差异可能很大。从现代的生物数据中得出的一个重要的见解是:我们需要考虑物种在多个进化谱系上的嵌套。换句话说,我们对生物多样性的定义需要包括整个生命之树。
因此,生物多样性保护的决策需要考虑整个生命之树,而不仅仅是单个物种,并在景观背景下考虑所有这些多样性。为了实现这一点,一个名为“空间系统发育学”的新领域被开发出来,该领域基本上将生命之树变成了一个GIS层。这种方法结合了两个主要元素,一个是系统发育,另一个是生物地理分布的空间数据集。多样性和特有性是根据系统发育相关性(即生物体与共同祖先的亲缘关系)、分支或谱系长度(例如从共同祖先推断出的遗传变化)以及系统发育中每个谱系的地理范围大小来衡量的。这种定义生物多样性的进化方法极大地推动了生态学和进化中许多问题的研究,并提供了实用的定量指标来促进保护和管理决策。
基于上述考虑,我们不能假设一种与另一种具有相同分类学名称的植物在任何特定方面都是相同的。例如,在修复过程中,我们不能仅仅因为苗圃中的植物与历史上存在于修复地点的植物具有相同的物种名称,就认为它们具有相同的特性或是很好的替代品。传统物种命名的谱系之间存在重要差异,这些差异往往与它们之间的生态或地理距离有关。而基因在更大的距离和生态空间内流动往往受到限制,当地植物种群往往通过适应当地环境和基因漂移发展出不同的属性。这是长期以来的论点的基础,即必须使用本地基因型进行恢复。

对加州植物进行的广泛的精细系统发育研究结果表明,隐蔽多样性在我们的植物群和加州自然景观中广泛存在(即,需要进行分类学识别的独特进化谱系,在形态学上很难或不可能区分)。考虑到加州植物多样性和特异性的异常水平,以及我们对环境异质性和加州自然景观的长期动态变化有利于植物谱系的进化和持久性的理解,这些发现并不令人惊讶。事实上,加利福尼亚植物的许多不同属和科都因其在形态学上难以区分而闻名,正如在年轻、积极进化的分支中所预期的那样,隐形谱系代表着不可替代的多样性,在生态上可能无法与其他此类谱系互换,在保护规划中至关重要。
不幸的是,很少有系统发育研究包括足够的种群样本,以确定加州植物群中以前未被发现或未被识别的植物进化谱系。即使是最仔细的研究也可能遗漏了现在可行的全基因组研究可能揭示的重要谱系。此外,已经进行的为数不多的详细研究的结果表明,很难可靠地预测自然景观中可能存在的进化或遗传多样性。这种困难甚至适用于具有独特历史和特征的密切相关的植物谱系。然而,跨生态和地理变异轴采样的策略对于发现隐藏的多样性很有用。
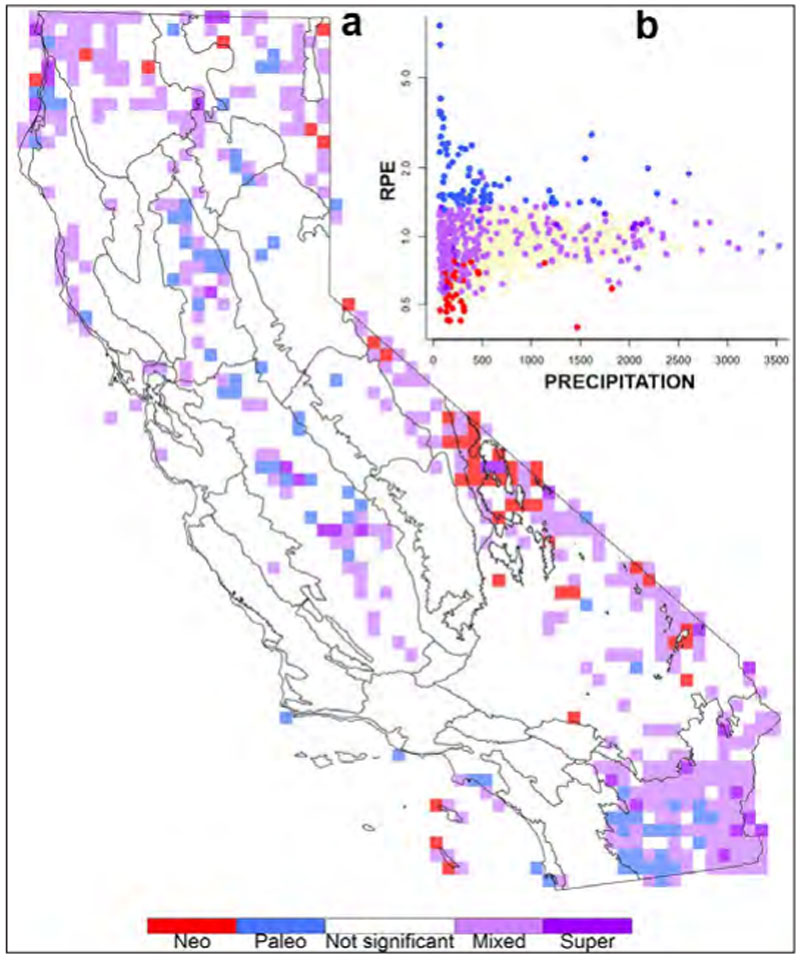
已命名物种内相当多的小尺度变异与植物的地理和生态环境有关。因此,如果缺乏系统发育研究,地理和生态距离可以作为亲缘关系的合理解释。此外,地理和生态环境在影响生物多样性的进化过程中发挥着积极作用;植物通常在地理上是局部适应的,并且分布在一个区域内的不同地点。出于这些原因,保护生态地理上不同的种群是很重要的,即使它们没有经过系统发育研究或没有分类学名称。同样重要的是,避免将生态地理上遥远的种群中的植物引入居民种群,因为这有稀释当地适应能力或危及当地特有种的风险。
这些发现和知识上的局限,加强了在其生态和地理范围内保护物种的重要性,并在进行恢复或其他涉及主动播种的保护行动时,尽可能使用最当地和采样最充分的种子来源。迄今为止,大多数关于植物种群在其范围内损失的担忧,以及当地获得的种子对恢复工作的重要性都是基于种群遗传因素,例如引入非本地植物可能导致本地适应性丧失和远缘繁殖抑制(即远亲杂交后代的适应性降低)。尽管这些问题至关重要,但假设目前的植物分类准确地反映了进化谱系的真实多样性,从而保证了分类识别。一个所谓的物种可能包括值得分类学承认的多个进化谱系,这些谱系可能由于杂交和基因淹没、繁殖干扰或引入的相关物种的竞争而灭绝。
关于隐性多样性的另一个未被充分重视的问题是,以野生植物谱系为代表的大量未开发的生物数据仍有待充分采样和理解。我们正处于大规模负担得起的全基因组测序和强大的基因组分析新方法的前沿,这些方法可能会彻底改变我们对植物进化、生态和生物地理历史的理解,包括对年轻和古老谱系的发现或重新解释。涉及在全州范围内广泛重新分配恢复种质资源,或通过基因增强方法,会很容易地消除或无可挽回地混淆这种野生进化信号,如果这种信号仍然可用,可能对明智地应对气候变化和其他人为对植物群的影响至关重要。在这个全球生物多样性热点地区,我们有可能失去一个发现和获取大量植物多样性知识的重要机会。出于所有这些原因,在保护工作中使用非本地种子应该是最后的策略。使用时,应在此类信息的中央存储库中对其进行跟踪和记录,记录人类发放种子的引入位置以及来源。这些信息可以极大地有助于从未来的野外收集中挽救历史遗传信号,将注意力集中在与易位地点相对隔离的种群上,或者解释被人类植物运动混淆的遗传数据。
我们都关心气候变化对加州植物群的影响,人们很自然地倾向于将植物转移到当前范围之外的假设的安全地点。会有许多善意的、基于保护的建议来移动植物,以缓解气候变化。然而,人为的迁徙或其他涉及重新分配植物的努力,需要我们对情况有更多的了解。例如,不同的研究表明,植物对气候变化的反应是独特而复杂的,并且贫瘠土壤上的植物可能更容易受到气候变化的影响,因为贫瘠土壤中有相当大比例的地方病。

另一个值得关注的问题是气候变化预测的规模和避难所的潜力,粗粒度研究可能会忽视这些问题。人类对植物的移动没有良好的记录,本地植物种群的适应性和扩散潜力也可能比通常人为的更大。例如,给定位置内的微环境变化可能在生态和进化上具有显著意义,这意味着一些微站点中已经存在的基因型可能能够耐受新出现的环境条件。另一个考虑因素是生态释放或竞争性释放,即在没有竞争对手或其他新的生态条件下,在田间植物之间通常看不到的变化。这是一种记录在案的现象,已在加州拉斯特尼亚进行了实验证明。换句话说,在自然选择作用于幼苗之后,现有遗传变异可能大于在普通田间条件下观察到的植物的估计值。植物在应对人为气候变化时有效分散的能力也可能被低估。例如,通过对冰川后景观的历史生物地理学研究的深入了解表明,动物介导的植物扩散和定植速度比以前认为的更快。
本土植物种群栖息地破碎化和种群的大量减少,以及加州一些景观缺乏环境异质性,无疑将给本土植物在气候变化下的生存带来挑战,并可能需要采取积极的保护干预措施。在某些可怕的情况下,辅助迁移或基因扩增可能是必要的,但只能作为有可靠科学依据的最后措施。其他干预措施,如战略性地移除非本土植物,虽然会消耗大量资源,但对恢复本土植物种群具有巨大潜力,例如在伯克利山天际线地区。
与在原生范围之外重新分配植物相比,这些干预措施对原生植物造成意外负面影响的可能性要小得多。在考虑可能导致不可替代的自然多样性和加州生物多样性宝贵知识丧失的不可逆转的行动时,预防原则总是必要的。
土地管理者和恢复主义者可能会面临清理场地,来建立新的植物群落或稀有植物种群的缓解措施。稀有植物种群可能由于栖息地的丧失和连通性的降低而降低了遗传变异。那么,我们如何权衡不作为的与不完美的行动或基于不完整知识的行动的风险呢?在尝试使用当地合适的材料种植时,我们现在可以做些什么来解决常见的障碍?

人类主导的选择和扩散的案例揭示了恢复种植的好处和缺点。对稀有植物的特殊考虑——可能的近亲繁殖与远亲繁殖抑制,由于栖息地破坏而减少的个体和种群数量,以及建立或重建种群的需要或愿望——需要具体的研究和理解才能适当解决。为恢复而收集和扩增野生种群必须遵循最佳实践,以支持而不是淹没当地的适应基因,并确保保护隐藏基因的多样性。
由于气候变化、入侵物种和土地利用变化等人为压力,我们的时间变得越来越紧迫。操纵自然系统、改善或修复它们的诱惑早已存在。我们在大规模范围内这样做的能力从未如此强大,但我们对潜在影响的理解也更好。尽管如此,对新影响的理解仍在不断涌现,例如加州生态恢复工作中广泛引入的入侵性植物病原体(疫霉菌)。我们可以看看20世纪初盛行的湿地排水和填水,或者20世纪30年代和40年代猖獗的植树活动,并看到今天海平面上升减缓和猖獗的植树的后果。成为别人的坏榜样是我们应该努力避免的,并将其纳入我们对恢复或任何其他本土植物运动的考虑中。






发表回复